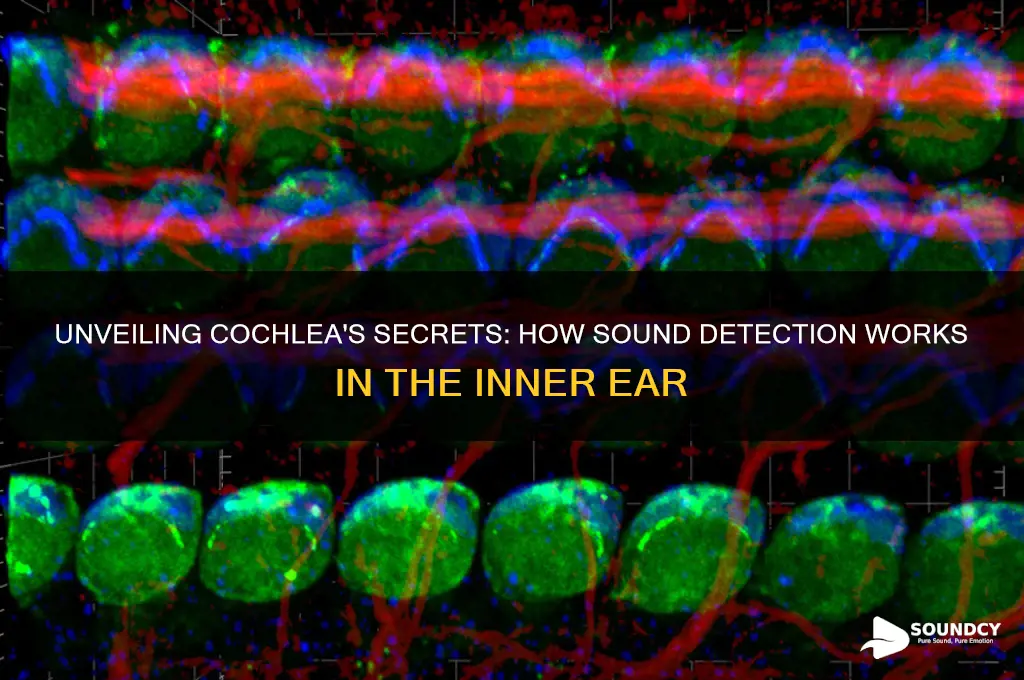
The detection of sound in the cochlea, a spiral-shaped organ in the inner ear, is a fascinating process that underpins our ability to hear. Sound waves enter the ear and travel through the auditory canal, causing the eardrum to vibrate. These vibrations are then transmitted to the cochlea via tiny bones in the middle ear, where they are converted into fluid motion. Within the cochlea, thousands of hair cells, each with stereocilia of varying lengths, are embedded in a gelatinous membrane. As the fluid moves, the stereocilia bend, triggering mechanical signals that are transduced into electrical impulses. These impulses are then transmitted via the auditory nerve to the brain, where they are interpreted as sound. This intricate mechanism allows the cochlea to detect a wide range of frequencies, enabling us to perceive the complexity of the auditory world.
| Characteristics | Values |
|---|---|
| Mechanism | Sound detection in the cochlea relies on the mechano-electrical transduction process. |
| Key Structure | Organ of Corti, containing hair cells (inner and outer) and supporting cells. |
| Hair Cells | Inner hair cells (IHCs) primarily transmit auditory information; outer hair cells (OHCs) amplify sound. |
| Stereocilia | Hair cells have stereocilia (hair-like projections) arranged in a staircase pattern, which bend in response to sound waves. |
| Transduction Process | Mechanical vibrations from sound waves cause stereocilia to deflect, opening ion channels (e.g., mechanotransduction channels like TMC1/TMC2), leading to depolarization of hair cells. |
| Basilar Membrane | Vibrates in response to sound, with different frequencies causing maximal vibration at specific locations along its length (tonotopy). |
| Tectorial Membrane | Overlies the stereocilia and moves relative to the basilar membrane, causing stereocilia deflection. |
| Neural Transmission | Hair cell depolarization releases neurotransmitters (e.g., glutamate) to spiral ganglion neurons, which transmit signals to the auditory nerve. |
| Frequency Discrimination | Achieved through the tonotopic organization of the basilar membrane and cochlea. |
| Amplification | Outer hair cells use electromotility (via prestin protein) to amplify sound vibrations. |
| Dynamic Range | The cochlea can detect sounds ranging from ~0 dB (threshold of hearing) to ~120 dB (threshold of pain). |
| Adaptation | Hair cells adapt to sustained stimuli, allowing continuous detection of changing sounds. |
| Damage Susceptibility | Hair cells are vulnerable to damage from loud noise, ototoxic drugs, and aging, leading to hearing loss. |
| Regeneration | Unlike birds and reptiles, mammalian hair cells do not regenerate once damaged. |
| Research Advances | Gene therapies and stem cell research are exploring ways to restore hair cell function. |
Explore related products






What You'll Learn
- Hair Cell Mechanotransduction: How hair cells in the cochlea convert sound vibrations into electrical signals
- Basilar Membrane Role: Frequency discrimination via basilar membrane’s place-specific vibrations in the cochlea
- Cochlear Amplification: Active processes enhancing sensitivity and frequency selectivity in sound detection
- Auditory Nerve Signaling: Transmission of electrical signals from hair cells to the brain via nerves
- Outer Hair Cell Function: Role in fine-tuning frequency sensitivity and cochlear mechanics
![]()
Hair Cell Mechanotransduction: How hair cells in the cochlea convert sound vibrations into electrical signals
The process of hearing begins when sound waves travel through the ear canal and reach the cochlea, a spiral-shaped organ in the inner ear. Within the cochlea, specialized sensory cells called hair cells play a crucial role in converting sound vibrations into electrical signals that the brain can interpret. Hair cell mechanotransduction is the intricate process by which these cells detect and respond to mechanical stimuli, such as sound-induced vibrations. This mechanism is fundamental to our sense of hearing, and its understanding has been a significant focus in auditory research.
Hair cells are named for their distinctive hair-like projections called stereocilia, which are arranged in bundles on the cell's apical surface. These stereocilia are embedded in a gelatinous membrane called the tectorial membrane, which moves in response to sound waves. When sound vibrations reach the cochlea, they cause the fluid within it to oscillate, setting the tectorial membrane and stereocilia in motion. The key to mechanotransduction lies in the precise arrangement and movement of these stereocilia. Each bundle is organized in a staircase-like pattern, with stereocilia of varying heights. When the bundle is deflected, the stereocilia move relative to each other, causing a shearing motion.
At the core of mechanotransduction are mechanosensitive ion channels located at the tips of the stereocilia. These channels are believed to be formed by transmembrane proteins, with the most prominent candidates being the transmembrane channel-like (TMC) proteins. When the stereocilia are deflected, the tension or displacement is transduced into a mechanical force that opens these ion channels. This allows specific ions, primarily potassium and calcium, to flow into the hair cell. The influx of positive ions depolarizes the cell, creating an electrical signal. This process is remarkably fast and sensitive, enabling hair cells to detect a wide range of sound frequencies and intensities.
The conversion of mechanical energy into electrical signals involves a complex interplay of various proteins and cellular structures. The stereocilia are connected by tip links, which are protein filaments composed of protocadherin 15 and cadherin 23. These tip links are thought to gate the mechanosensitive ion channels by transmitting the mechanical force from the moving stereocilia to the channel proteins. Additionally, the adaptation motor, a complex of proteins including myosin-1c, ensures that the hair cell can quickly reset and respond to continuous stimulation. This motor adjusts the tension on the tip links, allowing the hair cell to maintain its sensitivity over a wide dynamic range.
In summary, hair cell mechanotransduction is a sophisticated process that transforms sound-induced vibrations into electrical impulses. The precise arrangement of stereocilia, the gating of ion channels, and the intricate protein interactions all contribute to the remarkable sensitivity and speed of this system. Understanding these mechanisms not only provides insights into the biology of hearing but also has implications for developing treatments for hearing loss and other auditory disorders. Research in this field continues to unveil the intricate details of how we perceive the world of sound.
How Does That Sound to You? Exploring the Impact of Tone and Delivery
You may want to see also
Explore related products






![]()
Basilar Membrane Role: Frequency discrimination via basilar membrane’s place-specific vibrations in the cochlea
The detection of sound in the cochlea is a complex process that relies heavily on the basilar membrane, a specialized structure within the cochlea. This membrane plays a pivotal role in frequency discrimination, enabling the auditory system to distinguish between different sound frequencies. When sound waves enter the ear, they travel through the auditory canal and cause the eardrum to vibrate. These vibrations are then transmitted to the cochlea via the ossicles, a series of tiny bones in the middle ear. Upon reaching the cochlea, the vibrations interact with the basilar membrane, initiating a series of events that ultimately lead to the perception of sound.
The basilar membrane is unique in its structure, being wider and more flexible at the apex (beginning) and narrower and stiffer at the base (end). This tonotopic organization is crucial for frequency discrimination, as it allows different regions of the membrane to vibrate in response to specific frequencies. High-frequency sounds (e.g., high-pitched noises) cause the basilar membrane to vibrate more vigorously near the base, while low-frequency sounds (e.g., deep tones) elicit stronger vibrations near the apex. This place-specific vibration pattern is fundamental to the cochlea's ability to analyze and encode sound frequencies.
As the basilar membrane vibrates, it stimulates the hair cells that sit atop it. These hair cells, both inner and outer, are equipped with stereocilia—microscopic hair-like projections that bend in response to the membrane's movement. The bending of stereocilia opens ion channels, triggering an electrical signal that is transmitted to the auditory nerve. The precise location of this stimulation along the basilar membrane corresponds to the frequency of the sound, a principle known as the "place theory" of hearing. This mechanism ensures that each frequency activates a specific region of the cochlea, allowing the brain to accurately interpret the pitch of the sound.
The role of the basilar membrane in frequency discrimination is further enhanced by its mechanical properties. The membrane's stiffness and mass vary along its length, creating a gradient that resonates with different frequencies. This resonance acts as a filter, amplifying certain frequencies while dampening others. For example, a 1000 Hz tone will cause maximal vibration at a specific location on the basilar membrane, while a 500 Hz tone will peak at a different, more apical location. This precise mapping of frequencies to places on the membrane is essential for the auditory system to discern the complex spectrum of sounds in our environment.
In summary, the basilar membrane is a critical component in the cochlea's ability to detect and discriminate sound frequencies. Its place-specific vibrations, combined with the tonotopic arrangement of hair cells, enable the auditory system to encode and interpret a wide range of frequencies. By translating sound waves into spatial patterns of activity along the membrane, the cochlea provides the foundation for our sense of hearing. Understanding the basilar membrane's role not only sheds light on the intricacies of auditory processing but also highlights the elegance of the biological mechanisms that underlie our perception of sound.
Sound Walls: Quieting the Highway Roar
You may want to see also
Explore related products






![]()
Cochlear Amplification: Active processes enhancing sensitivity and frequency selectivity in sound detection
The cochlea, a spiral-shaped organ in the inner ear, is responsible for converting sound vibrations into electrical signals that the brain can interpret. Cochlear amplification is a critical active process that enhances the sensitivity and frequency selectivity of sound detection, allowing us to perceive a wide range of frequencies and intensities. This process is facilitated by the outer hair cells (OHCs) in the organ of Corti, which are uniquely adapted to amplify and tune incoming sound signals. OHCs achieve this through a mechanism known as electromotility, where changes in their membrane potential cause rapid alterations in their cell length. This active movement amplifies the traveling wave along the basilar membrane, increasing the displacement of the stereocilia on both inner and outer hair cells, thereby boosting sensitivity to faint sounds.
Frequency selectivity, another key aspect of cochlear amplification, is achieved through the tonotopic organization of the basilar membrane and the active feedback provided by OHCs. The basilar membrane is graded in stiffness and width, with different regions resonating at specific frequencies. When sound enters the cochlea, it travels along the basilar membrane, causing maximal vibration at a specific location depending on its frequency. OHCs further refine this process by actively reducing the width of the frequency tuning curve, a phenomenon known as "sharpening." This is accomplished through the force generated by OHC electromotility, which feeds energy back into the system, enhancing the precision with which frequencies are discriminated.
The active processes of OHCs are driven by the motor protein prestin, which is essential for electromotility. Prestin undergoes conformational changes in response to voltage fluctuations, enabling OHCs to contract and expand rapidly. This mechanism not only amplifies the signal but also operates within a specific frequency range, contributing to the cochlea's ability to distinguish between closely spaced frequencies. The nonlinear behavior of prestin ensures that amplification is most effective at low sound levels, preserving the dynamic range of hearing and preventing saturation at higher intensities.
Cochlear amplification also involves a delicate balance between active and passive mechanisms. While OHCs provide the active component, the mechanical properties of the basilar membrane and the tectorial membrane contribute passively to frequency selectivity. The interaction between these elements ensures that the cochlea can detect sounds across a vast intensity range (from whispers to loud noises) while maintaining sharp frequency discrimination. Dysfunction of OHCs, as seen in conditions like age-related hearing loss or noise-induced hearing damage, leads to reduced amplification and degraded frequency selectivity, highlighting the importance of these active processes in normal hearing.
In summary, cochlear amplification is a sophisticated active process that relies on the electromotility of OHCs to enhance sensitivity and frequency selectivity in sound detection. By amplifying mechanical signals and sharpening frequency tuning, this mechanism enables the cochlea to perform its dual role of detecting faint sounds and distinguishing between complex auditory stimuli. Understanding these processes not only sheds light on the remarkable capabilities of the auditory system but also informs the development of treatments for hearing impairments that arise from the dysfunction of cochlear amplification.
Exploring the Rich, Deep, and Versatile Sound of the Double Bass
You may want to see also
Explore related products






![]()
Auditory Nerve Signaling: Transmission of electrical signals from hair cells to the brain via nerves
The process of auditory nerve signaling is a complex yet fascinating mechanism that enables us to perceive sound. In the cochlea, sound detection begins with the vibration of the basilar membrane, which is lined with specialized sensory cells called hair cells. These hair cells are equipped with stereocilia, tiny hair-like projections that bend in response to the mechanical vibrations caused by sound waves. When sound enters the cochlea, it triggers the movement of the basilar membrane, causing the stereocilia to deflect. This deflection initiates a series of events that ultimately lead to the transmission of electrical signals to the brain.
As the stereocilia bend, they open ion channels in the hair cell membrane, allowing ions such as potassium and calcium to flow into the cell. This influx of ions creates an electrical signal, known as a receptor potential, which is then transmitted to the base of the hair cell. At the base, the hair cell synapses with auditory nerve fibers, forming a specialized junction called a ribbon synapse. Here, the electrical signal is converted into a chemical signal through the release of neurotransmitters, primarily glutamate, into the synaptic cleft. The neurotransmitters bind to receptors on the auditory nerve fibers, triggering an action potential that propagates along the nerve.
The auditory nerve fibers, also known as afferent neurons, carry these action potentials from the cochlea to the brainstem. This transmission occurs via the eighth cranial nerve, which is composed of thousands of individual nerve fibers, each responding to a specific frequency range. The action potentials travel rapidly along the auditory nerve, ensuring that the timing and intensity of the original sound are preserved. The nerve fibers are myelinated, which increases the speed of signal conduction, allowing for precise temporal coding of sound information.
Once the signals reach the brainstem, they are processed in the cochlear nucleus, the first relay station for auditory information. Here, the signals are sorted, filtered, and transmitted to higher auditory centers in the brain, including the superior olivary complex, inferior colliculus, and eventually, the auditory cortex. Each of these regions plays a crucial role in decoding the complex patterns of neural activity, enabling us to perceive and interpret sound. The auditory cortex, in particular, is responsible for higher-order processing, such as sound recognition, localization, and integration with other sensory inputs.
The efficiency and precision of auditory nerve signaling are remarkable, allowing us to detect a wide range of sound frequencies, intensities, and temporal patterns. This process relies on the intricate interplay between the mechanical vibrations of the cochlea, the transduction of these vibrations into electrical signals by hair cells, and the rapid transmission of these signals to the brain via the auditory nerve. Damage to any component of this pathway, such as hair cell loss or nerve degeneration, can lead to hearing impairment or deafness, underscoring the critical importance of each step in the auditory signaling process. Understanding these mechanisms not only sheds light on the complexity of hearing but also informs the development of treatments and technologies to address hearing disorders.
The Power of Sound: Enhancing Human Health, Mind, and Well-being
You may want to see also
Explore related products






![]()
Outer Hair Cell Function: Role in fine-tuning frequency sensitivity and cochlear mechanics
The detection of sound in the cochlea is a complex process that relies on the intricate interplay of various cellular and mechanical components. Among these, outer hair cells (OHCs) play a pivotal role in fine-tuning frequency sensitivity and optimizing cochlear mechanics. OHCs are specialized sensory cells located within the organ of Corti, and their unique electromotile properties enable them to amplify and refine the mechanical vibrations induced by sound waves. This active process, often referred to as the cochlear amplifier, is essential for enhancing the sensitivity, frequency selectivity, and dynamic range of hearing.
Outer hair cells achieve their function through a mechanism known as somatic electromotility, which allows them to change their length in response to electrical signals. When sound waves travel through the cochlea, they cause the basilar membrane to vibrate, deflecting the stereocilia of both inner and outer hair cells. This deflection opens mechanically gated ion channels, primarily allowing potassium ions to flow into the cell. The resulting change in membrane potential triggers OHCs to rapidly contract or elongate, a process driven by the motor protein prestin located in their lateral membrane. This electromotile response amplifies the basilar membrane's vibrations, particularly at low sound levels, thereby increasing the sensitivity of the cochlea.
The role of OHCs in fine-tuning frequency sensitivity is closely tied to their tonotopic arrangement along the basilar membrane. Different regions of the basilar membrane are tuned to specific frequencies, with high frequencies detected at the base and low frequencies at the apex. OHCs in each region are specialized to amplify vibrations within their respective frequency range. This spatial organization, combined with the active feedback provided by OHC electromotility, sharpens the frequency selectivity of the cochlea, allowing for precise discrimination of sound frequencies.
In addition to amplifying sound, OHCs contribute to cochlear mechanics by maintaining the proper stiffness and damping of the organ of Corti. Their motility helps to dampen excessive vibrations, preventing overstimulation and potential damage to the delicate sensory structures. Furthermore, OHCs are involved in the regulation of cochlear fluid dynamics, ensuring that the endolymph and perilymph compartments function optimally to transmit sound-induced vibrations. This dual role in both amplification and mechanical stabilization underscores the importance of OHCs in maintaining the integrity of auditory transduction.
The dysfunction or loss of outer hair cells, often observed in age-related hearing loss or noise-induced hearing damage, leads to a significant reduction in cochlear amplification and frequency selectivity. This results in decreased hearing sensitivity, particularly for soft sounds, and impaired ability to distinguish between closely spaced frequencies. Understanding the precise mechanisms by which OHCs fine-tune frequency sensitivity and cochlear mechanics is crucial for developing therapeutic strategies to mitigate hearing impairments and restore auditory function. In summary, outer hair cells are indispensable for the cochlea's ability to detect and discriminate sounds with remarkable precision, making them a focal point in auditory research and clinical interventions.
Do Eclipse Curtains Block Sound?
You may want to see also
Frequently asked questions
The primary method is through the mechanical vibration of hair cells in the organ of Corti, which are stimulated by the movement of fluid within the cochlea.
Hair cells have stereocilia (tiny hair-like projections) that bend in response to fluid movement, opening ion channels and generating electrical signals that are transmitted to the auditory nerve.
The basilar membrane vibrates at different frequencies along its length, allowing specific regions to respond to specific sound frequencies, a process known as tonotopy.
Yes, damage to the hair cells or other structures in the cochlea can lead to hearing loss, as the ability to convert sound waves into electrical signals is compromised.































