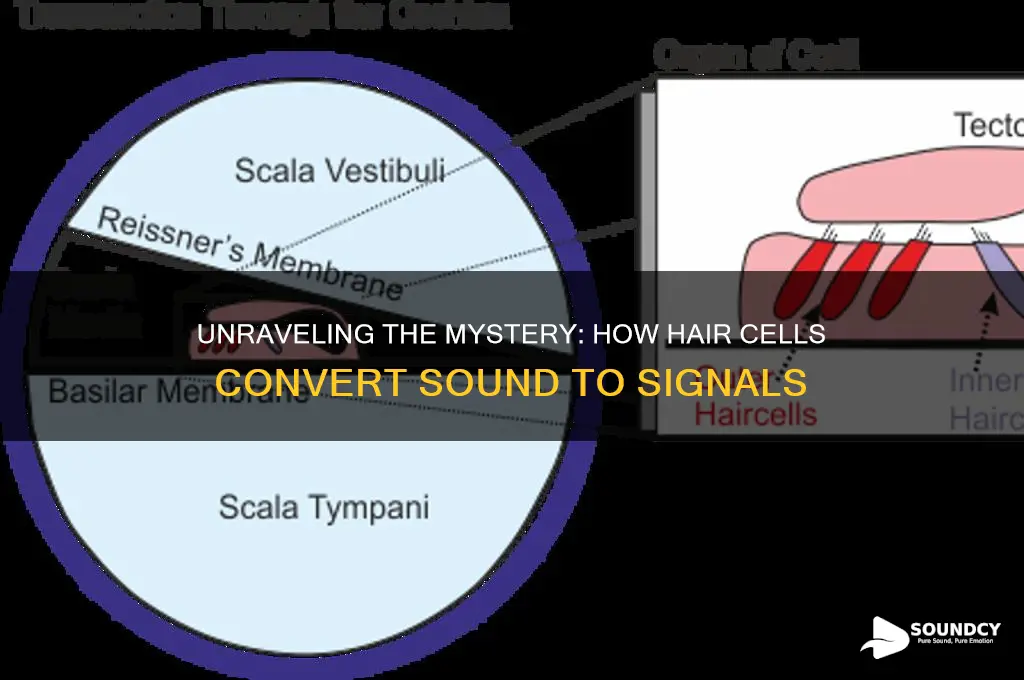
Hair cells, specialized sensory cells located within the cochlea of the inner ear, play a crucial role in the transduction of sound waves into electrical signals that the brain can interpret. These cells are characterized by their apical stereocilia, which are arranged in a staircase-like pattern and are embedded in a gelatinous membrane called the tectorial membrane. When sound waves travel through the cochlea, they cause the basilar membrane to vibrate, which in turn displaces the stereocilia. This mechanical displacement triggers the opening of ion channels at the tips of the stereocilia, allowing ions such as potassium and calcium to flow into the cell. The influx of ions changes the hair cell's membrane potential, generating an electrical signal that is transmitted to the auditory nerve fibers. This process, known as mechanotransduction, converts the physical energy of sound into neural signals, enabling the perception of sound.
Explore related products






What You'll Learn
- Mechanical Stimulation of Stereocilia: Sound waves cause stereocilia to bend, initiating transduction
- Ion Channel Opening: Tip links pull, opening mechanotransduction channels for ion influx
- Hair Cell Depolarization: Ion influx creates receptor potential, leading to cell depolarization
- Synaptic Transmission: Depolarization triggers neurotransmitter release to auditory nerve fibers
- Adaptation Mechanisms: Hair cells reset sensitivity via myosin motors and calcium feedback
![]()
Mechanical Stimulation of Stereocilia: Sound waves cause stereocilia to bend, initiating transduction
The process of sound transduction in the inner ear begins with the mechanical stimulation of stereocilia, the microscopic hair-like structures atop hair cells in the organ of Corti. When sound waves travel through the cochlea, they cause the basilar membrane to vibrate. This vibration is frequency-specific, meaning different regions of the basilar membrane respond to different sound frequencies. As the basilar membrane moves, it deflects the stereocilia, which are embedded in a gelatinous structure called the tectorial membrane. This deflection is the critical first step in converting mechanical energy into electrical signals that the brain can interpret as sound.
Stereocilia are organized in rows of increasing height, resembling a staircase, and are interconnected by tip links—protein filaments composed of protocadherin 15. When sound waves cause the stereocilia to bend, either toward the tallest stereocilium (positive deflection) or away from it (negative deflection), the tip links act as gating springs. In the case of positive deflection, the tension on the tip links opens mechanotransduction channels located at the tips of the stereocilia. These channels allow positively charged ions, primarily potassium (K⁺) and calcium (Ca²⁺), to flow into the hair cell from the endolymph, a potassium-rich fluid surrounding the hair cells.
The influx of positive ions depolarizes the hair cell, creating an electrical signal known as a receptor potential. This depolarization triggers the release of neurotransmitters at the base of the hair cell, which then transmit the signal to auditory nerve fibers. The specificity of this process ensures that different sound frequencies are encoded based on which region of the basilar membrane, and consequently which hair cells, are activated. Thus, mechanical stimulation of stereocilia is directly translated into neural activity.
It is important to note that the stereocilia are highly sensitive structures, capable of detecting deflections on the nanometer scale. This sensitivity is crucial for perceiving the wide range of sound intensities and frequencies that humans can hear. Additionally, the rapid adaptation of hair cells allows them to respond continuously to sustained sounds, ensuring that the auditory system remains dynamic and responsive. Without the precise mechanical stimulation of stereocilia, sound transduction would not occur, and auditory perception would be impossible.
In summary, the bending of stereocilia by sound-induced vibrations is the fundamental mechanism that initiates the transduction of mechanical sound energy into electrical signals. This process relies on the intricate structure of stereocilia, the role of tip links, and the opening of mechanotransduction channels. Understanding this mechanism provides insight into the remarkable ability of the auditory system to detect and interpret a vast array of sounds with precision and fidelity.
How Stone Matrix Asphalt Reduces Noise Pollution
You may want to see also
Explore related products






![]()
Ion Channel Opening: Tip links pull, opening mechanotransduction channels for ion influx
The process of sound transduction in hair cells begins with the intricate mechanism of ion channel opening, specifically through the action of tip links. Hair cells, located in the organ of Corti within the cochlea, possess stereocilia—stiff, hair-like projections arranged in bundles of increasing height. At the tips of these stereocilia are protein filaments called tip links, which connect the top of each stereocilium to the side of its taller neighbor. When sound waves travel through the cochlear fluid, they cause the stereocilia to deflect, exerting tension on these tip links. This mechanical force is the critical first step in converting sound into electrical signals.
The tip links are not merely structural components; they are directly coupled to mechanotransduction channels embedded in the stereocilia's plasma membrane. These channels are highly specialized proteins that remain closed until the tip links experience sufficient tension. When sound-induced deflection pulls on the tip links, they act as a molecular tether, transmitting the mechanical force to the mechanotransduction channels. This force causes the channels to open, allowing ions from the surrounding fluid—primarily potassium (K⁺) and calcium (Ca²⁺)—to rush into the hair cell. This influx of positively charged ions depolarizes the cell, generating an electrical signal that the hair cell can then transmit to the auditory nerve.
The opening of these mechanotransduction channels is rapid and highly sensitive, enabling hair cells to detect even the faintest sounds. The channels are thought to be composed of transmembrane proteins such as TMC1 and TMC2, which form the pore through which ions flow. The precise mechanism by which tip link tension gates these channels remains a topic of research, but it is clear that the tip links play a pivotal role in translating mechanical energy into electrical activity. Without this mechanism, the hair cell would be unable to transduce sound waves into neural signals that the brain can interpret.
Once the mechanotransduction channels open, the resulting ion influx triggers a cascade of events within the hair cell. The depolarization caused by the ion entry opens voltage-gated calcium channels, further amplifying the signal. This calcium influx, in turn, initiates the release of neurotransmitters at the hair cell's synaptic terminals, relaying the signal to auditory nerve fibers. The entire process is remarkably efficient, allowing for real-time encoding of sound frequency, intensity, and timing. After the stimulus ceases, the tip links relax, the mechanotransduction channels close, and the hair cell returns to its resting state, ready to respond to the next sound wave.
In summary, the opening of mechanotransduction channels via tip link tension is a fundamental step in hair cell sound transduction. This mechanism exemplifies the elegance of biological systems in converting physical stimuli into electrical signals. The sensitivity and speed of this process are essential for our ability to perceive a wide range of sounds, from whispers to loud noises. Understanding this mechanism not only sheds light on auditory physiology but also informs efforts to develop treatments for hearing loss caused by hair cell dysfunction.
Koorui Monitor: Does It Have Audio?
You may want to see also
Explore related products





![]()
Hair Cell Depolarization: Ion influx creates receptor potential, leading to cell depolarization
Hair cell depolarization is a critical process in the transduction of sound into electrical signals within the inner ear. This mechanism begins when sound waves cause the stereocilia—the hair-like projections on the apical surface of hair cells—to deflect. Stereocilia are organized in rows of increasing height, and their deflection occurs either toward or away from the tallest row, depending on the direction of the sound-induced movement. When stereocilia are deflected, mechanotransduction channels located at the tips of these structures open, allowing ions to flow into the cell. The endolymph, the fluid surrounding the hair cells, is rich in potassium ions (K⁺), which are the primary charge carriers in this process. The influx of K⁺ ions creates a receptor potential, which is the initial electrical change in the hair cell.
The receptor potential is directly tied to the opening of mechanotransduction channels. These channels are highly selective for K⁺ ions due to the unique composition of the endolymph. As K⁺ ions enter the cell, they shift the membrane potential toward the equilibrium potential for potassium (EK⁺), which is typically positive relative to the resting potential. This shift results in depolarization of the hair cell, meaning the intracellular voltage becomes less negative. The magnitude of depolarization is proportional to the number of channels opened and the extent of stereocilia deflection, making the process highly sensitive to the intensity and frequency of the sound stimulus.
Depolarization of the hair cell triggers the release of neurotransmitters, such as glutamate, at the basal end of the cell. These neurotransmitters then activate afferent nerve fibers of the auditory nerve, transmitting the signal to the brain. The precision of this depolarization-driven signaling is essential for encoding the features of sound, including its amplitude and frequency. Importantly, hair cells also possess efferent synapses, which allow the brain to modulate their sensitivity and protect them from overstimulation.
The ion influx responsible for depolarization is not only rapid but also highly localized, ensuring that the electrical signal accurately reflects the mechanical input. This localization is maintained by the tight arrangement of stereocilia and the precise distribution of mechanotransduction channels. Additionally, the hair cell’s apical and basal compartments are electrically isolated, which enhances the efficiency of signal transduction. This compartmentalization ensures that the receptor potential generated at the stereocilia is effectively transmitted to the basolateral membrane, where neurotransmitter release occurs.
In summary, hair cell depolarization is a key step in auditory transduction, driven by the influx of K⁺ ions through mechanotransduction channels. This ion influx generates a receptor potential, leading to depolarization of the hair cell and subsequent neurotransmitter release. The process is finely tuned to capture the nuances of sound, relying on the precise arrangement of stereocilia and the unique ionic composition of the endolymph. Understanding this mechanism provides insights into how the ear converts mechanical energy into neural signals, forming the basis of hearing.
Cows and Ducks: Unique or Similar Calls?
You may want to see also
Explore related products






![]()
Synaptic Transmission: Depolarization triggers neurotransmitter release to auditory nerve fibers
The process of sound transduction in the auditory system is a fascinating mechanism that involves the intricate workings of hair cells within the cochlea. When sound waves reach the inner ear, they initiate a complex series of events, ultimately leading to the transmission of auditory information to the brain. This journey begins with the stimulation of hair cells, which are specialized sensory cells in the organ of Corti. These cells are equipped with stereocilia, hair-like projections of varying heights, forming a V-shaped bundle. When sound waves cause the basilar membrane to vibrate, the stereocilia move, triggering a mechanical response.
Depolarization and Synaptic Transmission: As the stereocilia deflect, ion channels open, allowing ions such as potassium and calcium to flow into the hair cell. This influx of ions leads to depolarization, a critical step in the transduction process. Depolarization is the process by which the hair cell's membrane potential becomes less negative, moving towards a positive value. This change in voltage is a key signal that initiates the next phase of auditory information processing. The depolarized hair cell now prepares to communicate with the auditory nerve fibers, a process known as synaptic transmission.
At the base of each hair cell, there are synaptic terminals containing vesicles filled with neurotransmitters, primarily glutamate. When depolarization occurs, these vesicles fuse with the cell membrane, releasing their neurotransmitter content into the synaptic cleft. This release is a highly regulated process, ensuring that the right amount of neurotransmitter is delivered at the appropriate time. Glutamate, as the primary neurotransmitter, plays a crucial role in exciting the auditory nerve fibers, also known as afferent neurons. These neurons have receptors that respond specifically to glutamate, ensuring a precise and efficient transmission of the auditory signal.
The released glutamate molecules bind to postsynaptic receptors on the auditory nerve fibers, triggering a new wave of depolarization in these neurons. This depolarization generates an action potential, an electrical signal that travels along the auditory nerve towards the brainstem. Each action potential carries information about the characteristics of the original sound wave, including its frequency and intensity. The precision of this synaptic transmission is vital, as it ensures that the brain receives an accurate representation of the auditory stimulus.
In summary, the depolarization of hair cells is a critical trigger for neurotransmitter release, facilitating communication with auditory nerve fibers. This synaptic transmission is a rapid and highly coordinated process, essential for converting mechanical sound energy into electrical signals that the brain can interpret. Understanding these mechanisms provides valuable insights into the remarkable ability of the auditory system to transduce sound, ultimately enabling our sense of hearing.
FLAC vs MP3: Comparing Audio Quality and Sound Differences
You may want to see also
Explore related products






![]()
Adaptation Mechanisms: Hair cells reset sensitivity via myosin motors and calcium feedback
Hair cells, the sensory receptors in the inner ear, play a crucial role in transducing sound into electrical signals that the brain can interpret. To maintain their sensitivity across a wide range of sound intensities and frequencies, hair cells employ adaptation mechanisms. These mechanisms ensure that hair cells can quickly reset their sensitivity after being stimulated, allowing them to respond effectively to continuous or changing auditory input. Central to this process are myosin motors and calcium feedback, which work in concert to restore the hair cell’s resting state.
Myosin motors, specifically myosin VI and myosin VIIa, are essential components of the adaptation process. These molecular motors are anchored to the stereocilia—the hair-like projections on the apical surface of hair cells—and interact with actin filaments within the stereocilia. When sound waves cause the stereocilia to deflect, mechanotransduction channels open, allowing ions such as potassium and calcium to flow into the cell. This influx triggers a cascade of events, including the activation of myosin motors. Myosin VI, in particular, is thought to move along actin filaments, pulling on the stereociliary links (tip links) that connect adjacent stereocilia. This action helps restore the resting position of the stereocilia, closing the mechanotransduction channels and halting the influx of ions.
Calcium feedback is another critical component of the adaptation mechanism. As calcium ions enter the hair cell through the open mechanotransduction channels, they bind to calcium-binding proteins, such as calmodulin. This binding initiates a feedback loop that reduces the sensitivity of the mechanotransduction channels. Calcium-bound calmodulin activates an enzyme called myosin light chain kinase (MLCK), which phosphorylates myosin motors, modulating their activity. Additionally, calcium ions directly interact with the mechanotransduction channels, reducing their open probability. This dual action of calcium—both through myosin modulation and direct channel regulation—ensures rapid and precise adaptation.
The interplay between myosin motors and calcium feedback is finely tuned to allow hair cells to adapt to both sustained and transient stimuli. For example, during prolonged sound exposure, calcium accumulation gradually decreases channel sensitivity, while myosin motors work continuously to reset the stereocilia. Conversely, for brief sounds, the adaptation mechanisms act swiftly to restore the hair cell’s resting state, preparing it for the next stimulus. This dynamic regulation is vital for the hair cell’s ability to encode a wide range of sound intensities and frequencies with high fidelity.
In summary, adaptation mechanisms in hair cells rely on the coordinated actions of myosin motors and calcium feedback to reset sensitivity after stimulation. Myosin motors physically reposition the stereocilia, while calcium ions modulate channel activity and motor function through feedback loops. Together, these processes ensure that hair cells remain responsive to auditory input, enabling the precise transduction of sound into neural signals. Understanding these mechanisms not only sheds light on the remarkable sensitivity and adaptability of the auditory system but also provides insights into potential therapeutic targets for hearing disorders.
Understanding Sound Amplitude: How Loud is Loud?
You may want to see also
Frequently asked questions
Hair cells, located in the cochlea of the inner ear, are specialized sensory cells responsible for converting mechanical sound vibrations into electrical signals. These signals are then transmitted to the brain via the auditory nerve, enabling hearing.
Hair cells have stereocilia, which are tiny hair-like projections on their apical surface. When sound waves cause the fluid in the cochlea to move, the stereocilia bend, triggering mechanical changes that open ion channels and initiate an electrical response.
Bending of the stereocilia causes tension on tip links, which connect adjacent stereocilia. This tension pulls open mechanotransduction channels, allowing ions like potassium and calcium to flow into the cell, depolarizing it and generating an electrical signal.
The cochlea is tonotopically organized, meaning different regions of the basilar membrane respond to specific frequencies. Hair cells in each region are tuned to detect particular frequencies based on their location along the membrane, allowing for frequency discrimination.































